Source: http://photobiology.info/Zhu.html
Xin-Guang Zhu
Plant Systems Biology Group, Institut za računalnu biologiju,
Kineska akademija znanosti/Max Planck društvo, Yue Yang Road,
Šangaj, Kina, 200031
www.picb.ac.cn/~zhuxinguang
Uvod
Zbog iscrpljivanja fosilnih goriva i rastuće potražnje za proizvodnjom hrane, biljni biolozi po prvi put u povijesti igraju važnu ulogu u rješavanju velikog izazova za društvo, tj. Identificiranje novih načina za povećanje produktivnosti biljke u promjenjivoj globalnoj klimi , Povećanje učinkovitosti pretvorbe fotosintetske energije jedna je od glavnih ruta koja nam je još uvijek dostupna za dramatično povećanje produktivnosti usjeva. Kombiniranje modela sustava s evolucijskim algoritmom daje jedan učinkovit i izvediv mehanizam za prepoznavanje novih pristupa inženjeru višoj fotosintezi.
Veći dio našeg znanja o fotosintezi dobiva se pomoću redukcionističkog pristupa, gdje je svaka pojedina komponenta sustava izolirana i proučavana samostalno. Ovim smo pristupom dobili detaljne biofizičke i biokemijske informacije o različitim komponentama fotosinteze. Istovremeno, moderna biotehnologija omogućava jednostavno podizanje ili spuštanje regulacije gotovo svakog gena u fotosintezi. Čak i uz sve te kapacitete i znanje, još uvijek smo daleko od mogućnosti da precizno inžinjimo fotosintetsku napravu za višu fotosintetsku brzinu. Ovo nije previše iznenađujuće s obzirom da je učinkovitost pretvorbe fotosinteze energije svojstvo sustava, tj. Ona se određuje interakcijom između različitih komponenti sustava, a koja nije određena bilo kojom pojedinom komponentom sustava.
S obzirom na to da je fotosinteza temelj za proizvodnju hrane i energije i povećanu potražnju kako u našem društvu, kako možemo inženjeriti višu fotosintezu? Drugim riječima, što ograničava fotosintezu za različite biljke u različitim sredinama? Odgovori na ta dva pitanja su ključ za povećanje produktivnosti biljaka. Odgovaranje na ova dva pitanja vrlo je zahtjevno. Više od stotinu proteina uključeno je u fotosintezu. Čak i ako je manipulacija ograničena na jednostavno mijenjanje količine proteina, umjesto njegovih svojstava, potencijalne permutacije će se pojaviti u milijunima.
S obzirom na to da se istodobno mogu stvoriti višestruki proteini, vrlo brzo postaje očito da je eksperimentalno podizanje ili smanjenje količine svakog proteina i ispitivanje njegovih utjecaja na produktivnost neučinkovito. Jedan potencijalni način rješavanja ovog problema je uzeti sustavski pristup. Pristup sustavu je razviti matematičke modele koji vjerno simuliraju sve biofizike i biokemijske procese koji su uključeni u fotosintezu, a zatim provode numeričke eksperimente kako bi identificirali ciljeve inženjeru za veću produktivnost.
Koristit æu model fotosintetskog metabolizma ugljika kako bi ilustrirala osnovnu metodologiju razvoja modela sustava i korištenjem evolucijskog algoritma kako bi se utvrdila optimalna raspodjela dušika u metabolizmu zbog veće fotosintetske brzine apsorpcije CO2. Također ću ukratko opisati i druge glavne primjene modela sustava fotosinteze, koji uključuju povezivanje genomskih podataka s fiziološkim podacima, proučavanje dinamičkih signala u fotosintezi, proučavanje značaja promjena u fotosintetskom aparatu radi povećanja fotosintetske učinkovitosti i radi kao osnova za ekofiziološki modeli.
Jedan od ključnih ciljeva istraživanja biološke biologije jest povećati fotosintezu
Tijekom posljednja četiri desetljeća, uzgoj biljaka je proizveo znatna povećanja potencijalnih prinosa mnogih usjeva. Potencijalni prinos definira se kao prinos koji genotip može postići bez biotskih i abiotičkih naprezanja. Prema načelima Monteita (1977), potencijalni prinos (Y) usjeva i primarne proizvodnje (Pn) na određenom mjestu određuje:
Pn = St.εi. εc/k …. (1a)
Y = η.Pn …. (1b)
Gdje je St je godišnji integral incidentnog sunčevog zračenja (MJ m-2), εi učinkovitost kojom zračenje presijeca usjev; εc učinkovitost kojom se presresti zračenje pretvara u biomasu; η indeks žetve ili učinkovitost kojom se biomasa razdjeljuje u žetveni proizvod; i k energetski sadržaj biljne mase (MJ g-1). Sv određuje mjesto i godina. k varira vrlo malo između vegetativnih organa, tipično u prosjeku 17 MJ/kg, iako zrna s značajnim udjelom ulja može imati veći energetski sadržaj (Roberts i sur., 1993). Pn je primarna produktivnost, tj. Ukupna biomasa biljke proizvedena tijekom vegetacijske sezone. Na temelju jednadžbe 1, Y se određuje kombiniranim proizvodom od tri učinkovitosti, od kojih svaka opisuje široka fiziološka svojstva usjeva: εi, εc i η. ε1 određuje brzina razvoja i zatvaranja krova, te dugovječnost, veličina i arhitektura nadstrešnica. εc je funkcija kombinirane fotosintetske brzine svih lišća unutar krova, manje gubitaka respiratornog usatka (Long et al., 2006).
U kontekstu jednadžbe 1, povećanje potencijalnog prinosa tijekom proteklih 40 godina rezultiralo je uglavnom povećanjem η i εi, dok su ostvareni prinosi poboljšani boljom oplodnjom i poboljšanom zaštitom bolesti, povećavajući time εi (recenzirani: Long et al., 2006). No, kako možemo dalje povećati Y? Zdravi usjevi suvremenih kultivara u optimiziranom razmaku presreli su najveći dio raspoloživog zračenja u svojoj vegetacijskoj sezoni, ograničavajući izglede za poboljšanje εi. Grain u suvremenim sortama žitarica može predstavljati 60% ukupne biomase tijekom žetve (Evan, 1993). Indeks žetve za zrno čini se da se približava gornjoj granici s obzirom da minimalna količina biomase mora ostati u biljnom tijelu, kako bi se osiguralo da vitalne hranjive tvari i rezerve mogu biti translocirane u zrnu i da računaju materijale stanične stijenke koje ne mogu biti degradiran. Indeks žetve za usjeve biomase još je veći. Ako se η i εi približavaju gornjoj granici, daljnji porast potencijalnog prinosa može se postići samo porastom εc, koji se određuje fotosintezom i disanjem.
Dakle, koja je teoretska maksimalna εc, i što je realizirana εc? Teoretska analiza sugerira da su maksimalna εc za C3 i C4 biljke 4,6% odnosno 6% (Slika 1, vidi pojedinosti izračuna u Zhu et al., 2008).
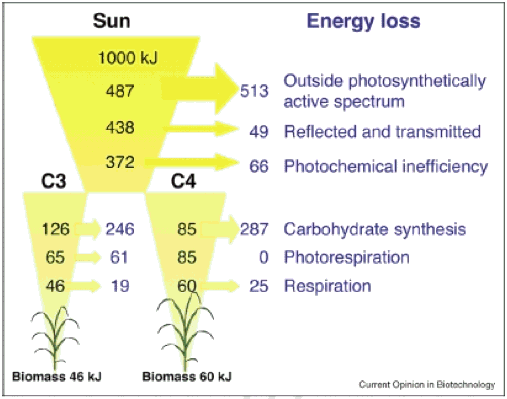
| Slika 1. Minimalni gubici energije izračunati za 1000 kJ incidentnog sunčevog zračenja, u svakom pojedinačnom koraku biljnog fotosintetskog procesa od presretanja zračenja do stvaranja pohranjene kemijske energije u biomasu. Razmatrane su i C3 i C4 (NADP-Malic Enzyme Type) fotosinteza. Izračuni pretpostavljaju temperaturu listova od 30° C i atmosferskog [CO2] od 380 ppm. Strelice pokazuju količinu gubitaka energije za različite procese (Zhu et al., 2008). |
Za C3 usjeve, najviša kratkoročna učinkovitost je oko 0,035, a za C4 oko 0,043; uobičajeno postizanje εc u polju je samo oko 1/3 od ovih maksimalnih vrijednosti, što upućuje na to da postoji znatna prostorija za daljnje povećanje εc (pregledano u Zhu et al., 2008). Izravni dokazi dali su biljke uzgojene u okviru obogaćivanja slobodnog zraka CO2, gdje su povećana i fotosintetska stopa i produktivnost biljke (Long et al., 2004).
Kako prepoznati nove načine da inženjeru za veću produktivnost? Kao što smo ranije spomenuli, eksperimentalni pristupi postaju neučinkoviti u identificiranju novih inženjerskih ciljeva. Ovdje ću upotrijebiti model fotosintetskog ugljikovog metabolizma kako bi ilustrirala sustavni pristup za prepoznavanje novih ciljeva za inženjering višu učinkovitost pretvorbe fotosintezne energije.
Biološki pristup sustavu kako bi se identificirale nove mogućnosti inženjer za veću produktivnost
U ovom odjeljku kratko ću opisati osnovni postupak za razvoj modela sustava, a zatim opisati metodu kombiniranja modela sustava s evolucijskim algoritmima kako bi se inženjeri otkrili novi ciljevi za veću produktivnost. Na kraju ovog odjeljka objasnit ću zašto evolucija nije odabrala optimalni fotosintetski aparat.
i) Osnovni postupak razvijanja modela sustava
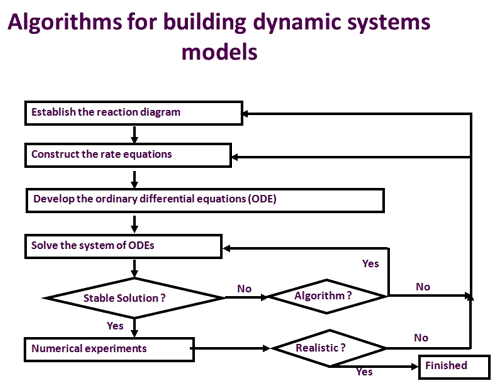
| Slika 2. Opći algoritam za izgradnju modela dinamičkih sustava. Proces uključen u razvoj modela sustava uključuje uspostavljanje dijagram reakcije, izgradnju jednadžbi stopa, izgradnju običnih diferencijalnih jednadžbi, testiranje i validaciju rezultata modela s eksperimentalnim opažanjima. Ovaj proces iterates sve dok model ne može vjerno reproducirati obično promatrane odgovore na genetske ili ekološke perturbacije. |
Slika 2 prikazuje proces koji se koristi za razvoj kinetičkog modela metaboličkog procesa. Prvo, reakcije koje su blisko uključene u ovaj proces potrebno je sastaviti kako bi se stvorio dijagram reakcije. Zatim se razvija jednadžba stope za svaku reakciju uključenu u dijagram. Model sustava koji predstavlja ovaj metabolički proces konstruira se razvijanjem obične diferencijalne jednadžbe (ODE) za svaki metabolit uključen u proces. Svaki ODE opisuje brzinu promjene koncentracije metabolita s vremenom, formiranim brzinama formiranja, umanjenoj stopi potrošnje tog metabolita. Na primjer, u fotosintetskom metabolizmu ugljika, RuBP (Ribuloza-1,5-bisfosfat) nastaje kroz fosforilaciju Ru5P (Ribuloza-5-fosfata) putem reakcije:
Ru5P+ATP→ ADP + RuBP …. 2
Upotrebljava se i pomoću RuBP karboksilacije i RuBP oksigenacije:
RuBP + CO2 → 2 PGA …. 3
RuBP + O2 → PGA + PGCA …. 4
gdje je PGA 3-fosfoglicerat i PGCA je 2-fosfoglikolat. Pod pretpostavkom da je brzina RuBP fosforilacije Vrp i stope RuBP karboksilacije (jednadžba 3) i oksigenacija (jednadžba 4) su Vc i Vo, tada se brzina promjene RuBP koncentracije s vremenom izražava kao:
d[RuBP]/dt = Vrp-Vc-Vo …. 5
Svi ODE kolektivno formiraju sustav običnih diferencijalnih jednadžbi, čije rješenje odgovara različitim fiziološkim i dinamičkim stanjima metaboličkog procesa.
Nakon što je konstruiran sustav običnih diferencijalnih jednadžbi, potreban je niz numeričkih eksperimenata za ispitivanje i potvrđivanje modela. Normalno, sustavni model mora pokazati svoju sposobnost dobivanja stalnih stanja, robusnosti prema vanjskoj perturbaciji i sposobnosti predviđanja uobičajenih fenomena. Na primjer, model fotosintetskog ugljikovog metabolizma dostigao je stabilno stanje u roku od oko 200 sekundi (Zhu et al., 2007). Vjerno je simulirao uobičajeno izmjerenu krivulju A-Ci, tj. Unos CO2 u odnosu na intercellularnu koncentraciju CO2 (Ci) krivulje, i za normalne (21%) i niske (2%) razine kisika. Nadalje, nakon drastične perturbacije međustanične koncentracije O2, model može brzo povratiti novo stabilno stanje s realnom brzinom fotosinteze. Konačno, model vjerno simulira fosfatno ograničenu fotosintezu u uvjetima niske stope upotrebe triosfosfata (Zhu et al., 2007). Nakon ovih validacija, model fotosintetskog metabolizma ugljika može se upotrijebiti za identifikaciju novih ciljeva za inženjersku višu fotosintetsku učinkovitost.
ii) Kombiniranje modela sustava s evolucijskim algoritmom za prepoznavanje novih ciljeva inženjeru za veću produktivnost
U tradicionalnom uzgoju, križevi između različitih biljaka stvaraju varijacije u određenim osobinama, poput prinosa usjeva. Sjeme za one osobe s vrhunskom kvalitetom osobina odabrano je i koristi se za generiranje pojedinaca za sljedeću generaciju. Nakon mnogih generacija, odabrane osobine postupno se poboljšavaju tijekom ovog procesa. Ova metoda dovela je do dramatičnog povećanja produktivnosti usjeva i stvorila “zelenu revoluciju”. Analogno odabiru boljih osobina tijekom generacija u tradicionalnom uzgoju, kombiniranje modela sustava s evolucijskim algoritmom omogućuje simuliranu evoluciju fotosintetskog aparata za veću brzinu fotosinteze.
Evolucijski algoritam u osnovi oponaša proces prirodnog odabira. U stvarnom prirodnom odabiru, selekcijski pritisak je osigurati preživljavanje i višu plodnost. U umjetnoj selekciji oponašan evolucijskim algoritmom, tlak selekcije je da ima veću brzinu fotosintetskog unosa CO2 s istom količinom dušikovog ulaganja u enzime u fotosintetskom ugljikovom metabolizmu. Nadalje, svaka generacija ima mnoge pojedince, pri čemu svaki pojedinac predstavlja jedan skup enzimskih koncentracija potrebnih za pokretanje modela metabolizma ugljika. Za svakog pojedinca, ili skup koncentracija enzima, postoji jedna stalna brzina apsorpcije CO2. U svakoj generaciji, pojedinac s višom stopom unosa fotosintetskih CO2 bit će odabran da “sjeme” sljedeće generacije. Kako bi generirali pojedince za sljedeće generacije, odabrana bi se osoba duplicirala kako bi generirala populaciju, pa će pojedinci u toj populaciji biti “mutirani” (tj. Dodavati slučajne varijacije koncentracijama enzima). Ovaj proces selekcije i mutacije iteratira, a brzina fotosintetskog udjela ugljičnog dioksida postepeno se povećava tijekom generacija. Ovaj proces prestaje kada brzina fotosinteze ne može biti povećana s više generacija (Zhu et al., 2007). Konačna distribucija dušika u skupini enzima smatra se optimalnom distribucijom dušika (Slika 3).
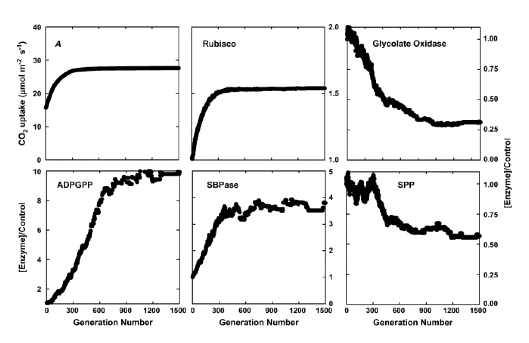
| Slika 3. Odabir optimalnog raspodjele resursa između enzima ugljikovog metabolizma kako bi se povećala svjetlosno zasićena fotosintetska brzina. A, povećava svjetlosno zasićenu fotosintetsku brzinu preko 1.500 generacija selekcije pomoću evolucijskog algoritma. B do F pokazuju promjene u količini od pet tih enzima u odnosu na njihove početne vrijednosti tijekom ovih 1.500 generacija. Selekcija je bila pri zasićenju svjetlosti, a Ci od 280 umola mol-1. |
Kratice:
A: fotosintetska stopa unosa CO2
ADPGPP: ADP glukoza pirofosforilaza
SBPaze: sedoheptuloza-l, 7-bisfosfataza
SPP: saharoza fosfat fosfataza
FBPaza: fruktoza-2,6-bisfosfataza.
Iz Zhu et al. (2007).
Kada se identificiraju optimalne distribucije dušika, možemo je koristiti za izračunavanje zahtjeva povećanja za svaki enzim,
Demand[i] = [Enzyme[i]_opt]/ [Enzyme[i]_initial] …. (6)
Kada [enzim [opt] predstavlja optimalnu koncentraciju dušika i [enzim [početni]] predstavlja početnu koncentraciju dušika. Ako je potražnja za enzimom veća od 1, aktivnost enzima treba povećati; U suprotnom, enzimska aktivnost mora biti ispuštena u slobodni dušik za sintezu drugih enzima kako bi se postigla optimalna stopa uzimanja CO2 (Zhu et al., 2007). Koristeći ovaj pristup, pokazali smo da Rubisco, SBPaze i FBPaze trebaju biti povećani za veću brzinu fotosintetskog CO2 (Zhu et al., 2007). Nadalje, povećanje kapaciteta sudopera, tj. Sposobnost konzumiranja triosfosfata (PGA, ili dihidroksiaceton fosfata ili gliceraldehid 3-fosfata) kroz sintezu škroba, saharoze ili drugih makromolekula. Predviđeno je da se dušik za povećanje aktivnosti ovih enzima prenosi iz fotorespirijskog puta (Zhu et al., 2007). SBPaze, jedan od enzima koji se utvrdi da se poveća za postizanje veće fotosinteze, prekomjerno je izražen u duhanu, što je dovelo do veće proizvodnje fotosinteze i proizvodnje biomase (Lefebvre et al., 2005), pokazujući da kombiniranje evolucijskog algoritma s modelima sustava može identificirati moguće inženjerske ciljeve za višu fotosintezu.
Drugi pristup za identifikaciju ciljeva inženjera je analiza metaboličke kontrole (MCA) pomoću izračuna koeficijenta kontrole protoka (Fell 1997). I MCA i evolucijski algoritam daju inženjerske ciljeve; međutim, postoje temeljne razlike između ova dva pristupa. Koeficijent kontrole protoka enzima predstavlja kontrolu tog enzima za određeni tok, uz pretpostavku da se sve druge razine enzima ne mijenjaju, tj. Taj koeficijent predstavlja lokalni pregled kontrole ovog enzima u odnosu na tok sustava. S druge strane, evolucijski algoritam identificira optimalnu raspodjelu svih enzima u metabolizmu istodobno mijenjajući sve koncentracije enzima. Stoga, teoretski, evolucijski algoritam može identificirati ciljeve za višu fotosintezu iz globalnijeg pogleda na sustav.
iii) Zašto fotosinteza, nakon milijuna godina evolucije, nije razvila optimalnu stopu fotosinteze?
Kombinacija evolucijskog algoritma s modelima sustava omogućava izvediv i učinkovit mehanizam za prepoznavanje optimalnog fotosintetskog aparata. Jedno filozofsko pitanje koje ljudi mogu postaviti je: nakon milijunima evolucije, zašto biljke nisu razvile optimalni fotosintetski aparat? Postoji nekoliko mogućih razloga. Prvo, evolucija odabire preživljavanje i plodnost, a ne produktivnost. Kako bi se osiguralo preživljavanje pod stresnim uvjetima, određena količina resursa, kao što je dušik, ulaže se u zaštitne mehanizme na štetu fotosintetske učinkovitosti. Na primjer, pokazali smo da je dušik pretjerano uložen u fotorespiratorni put (Zhu et al., 2007). Ova prevelika ulaganja, međutim, od presudne je važnosti za opstanak biljaka pod iznimno vrućim uvjetima. Iako su takvi uvjeti rijetki, možda jednom svakih nekoliko stotina godina, oni bi mogli biti dovoljno česti da potpuno eliminiraju biljke bez takvih zaštitnih mehanizama (Zhu et al., 2007). Drugo, globalna klimatska promjena je još jedan čimbenik. Koncentracija atmosferskog atmosferskog CO2 u posljednjih 25 milijuna godina bila je oko 280 ppm (Barnola et al., 1999), prije brzog porasta koncentracije CO2 nakon industrijske revolucije. Trenutni fotosintetski metabolizam ugljika, npr. Veća ulaganja u fotorespiratorni put, može predstavljati optimalnu raspodjelu dušika za razinu CO2 preindustrijske revolucije (Zhu et al., 2007). Prethodno smo pokazali da je jedan od kritičnih enzima u fotosintezi Rubisco doista optimiziran za veći unos CO2 pri koncentraciji CO2 od 280 ppm (Zhu et al., 2004). Dva suptilnija razloga također bi mogla biti odgovorna za trenutnu neoptimalnu raspodjelu dušika u fotosintetskom ugljikovom metabolizmu. Podoptimalno ulaganje SBPaze moglo bi biti povezano s ulogom SBPaze kao kontrolnom točkom u fotosintetskom ugljikovom metabolizmu; dok pretjerano ulaganje enzima u fotorespiratorni put također može biti posljedica strogog omjera RuBP karboksilacije i oksigenacije za određenu Rubisco specifičnost i atmosferske uvjete (Zhu et al., 2007).
Druge glavne primjene modela fotosintezi sustava
Modeli dinamičkih sustava imaju različite primjene osim identifikacije novih pristupa inženjeru za veću produktivnost. To uključuje:
Prvo, modeli sustava pružaju izravnu vezu genomskih podataka s fenotipskim podacima. Ulazi modela sustava su enzimske aktivnosti ili proteinski sadržaji, koji se mogu dobiti iz literaturne ankete ili proteomike. Izlazi modela sustava su koncentracije metabolita, tijekovi i fiziološki parametri.
Drugo, model sustava može se koristiti za proučavanje prilagodljivog značaja molekularnih promjena u fotosintetskom aparatu za fotosintetsku učinkovitost. Ovdje se fotosintetska aparatura definira kao različite komponente (enzimi, nosači prijenosa elektrona, itd.) I interakcije između različitih komponenata u fotosintezi. Kao što je ranije spomenuto, kombiniranje evolucijskih ili genetskih algoritama s modelom sustava omogućava učinkovit način prepoznavanja optimalnog fotosintetskog aparata pod definiranim uvjetima. Ova razlika između optimalnog fotosintetskog aparata i postojećeg fotosintetskog aparata omogućuje nam inženjer za veću produktivnost. Ova razlika također ukazuje na adaptabilnost značajnosti različitih svojstava fotosintetskog aparata.
Treće, modeli dinamičkih sustava mogu se koristiti za testiranje hipoteza o mehanizmima na kojima se temelje dinamički signali vezani uz fotosintezu. Upotrijebili smo model sustava primarnih događaja oko fotosustava II kako bismo proučili mehanističku osnovu indukcijske krivulje fluorescencije zabilježene nakon zasjenjenja zasićenog svjetlosnog impulsa (tj. Kratkog svjetlosnog impulsa (<1s) koji može zatvoriti sve reakcijske centre fotosinteze II) tamno prilagođeno lišće (Zhu et al., 2005).
Četvrto, modeli sustava fotosinteze mogu se koristiti u ekofiziološkim modelima simulacije usjeva kako bi se omogućilo izravno povezivanje genoma s ekosustavom. Računalno potpomognuto oblikovanje integriranog kruga (IC) odigralo je ključnu ulogu u brzom razvoju novih IC tehnologija. Očekuje se da će model koji može izravno povezati genom s ekosustavom može poslužiti istoj ulozi. Iako su modeli poljoprivrednih ekosustava razvijeni tijekom proteklih pola stoljeća, oni još uvijek ne mogu izravno predvidjeti učinkovitost usjeva iz genoma usjeva i njegovog okruženja rasta (Jones et al., 2003), iako postoji nekoliko nedavnih napora da se koriste QTL (tj. DNK koji su usko povezani s genima koji su podloga nekom svojstvu) u modelima agroekosustava. (Yin et al., 2004).
Razvijanje integriranog modela koji može izravno povezati genom s ekosustavom iznimno je zahtjevan jer takav model ne samo da mora mehanički opisati procese na različitim organizacijskim razinama, od razine ekosustava do razine kanape, listova i staničnih procesa, već također ima opisati promjene u ekspresiji gena pod različitim uvjetima i različitim razvojnim stadijima. Bez sumnje, razvoj takvog modela će trajati godinama zajedničkog napora cijele zajednice modeliranja biljaka. S obzirom da je fotosinteza osnova materijala i energije za rast i razvoj biljaka, detaljan mehanistički model fotosinteze je kritična komponenta takvog integriranog modela. Integracija modela fotosintetskog ugljikovog metabolizma s modelima agroekosustava mogla bi biti prvi korak prema realizaciji takvog integriranog modela genoma-ekosustava.
Literatura:
Barnola, JM, Raynaud, D., Lorius, C. Barkov, NI Povijesno COzapis iz Vostok ledene jezgre. U Trendovi: Zbornik Podaci o globalnih promjena. Ugljični dioksid analiza informacija centar, Oak Ridge National Laboratory, Zavod za energetska postrojenja, Oak Ridge, TN, SAD (1999).
Evans LT Crop Evolution, adaptacija i prinos. CUP, Cambridge. (1993)
Fell, DA Razumijevanje kontrolu metabolizma. Portland Press, London (1997).
Jones, JW, Hoogenboom, G., Porter, CH Boote, KJ, Batchelor, WD, Hunt, LA, Wilkens, PW, Singh, U., Gijsman, AJ, Ritchie, JT DSSAT modela obrezivanje sustava. European Journal of Agronomski 18, 235-265 (2003).
Lefebvre, S., Lawson, T., Zakhleniuk, OV, Lloyd, JC-Raines, CA Povećana sedoheptulose-1,7-bisfosfataze aktivnost u transgenskim biljkama duhana potiče fotosintezu i rast od rane faze u razvoju. Fiziologija bilja. 138, 451-460 (2005).
Duga, SP, Zhu, XG, Naidu, SL-Ort, poboljšanje DR Can fotosinteze povećati prinos usjeva? Plant Cell Environ. 29, 315-330 (2006).
Dugo SP, Ainsworth EA, Rogers A. i Ort DR Rising atmosferski ugljični dioksid: biljke lice svoju budućnost. Godišnji Recenzije Plant Biology, 55, 591-628 (2004).
Monteith, JL klime i učinkovitost proizvodnje usjeva u Velikoj Britaniji. Philos. Trans. R. Soc. London 281, 277-294 (1977).
Roberts MJ, Long SP, Tieszen LL i Beadle CL Mjerenje biljne biomase i neto primarne proizvodnje zeljasta raslinja. U: fotosinteze i proizvodnja u promjenjivoj okolini: Terenski i laboratorijski priručnik (EDS, učinite Hall, JMO Scurlock, HR Bolhàr-Nordenkampf, RC Leegood, a SP dugo), str 1-21.. Chapman & Hall, London (1993).
Yin, XY, Struik, računala i Kropff, MJ Uloga fiziologije bilja u predviđanju gen-u-fenotipa odnose. Trendovi Plant Sci. 9, 426-432 (2004).
Zhu, X.-G., Portis Jr., AR Long SP Bi transformacija C3 biljaka usjeva sa stranim Rubisco povećanje produktivnosti? Računalna analiza ekstrapolacije iz kinetičkih svojstava nadstrešnica fotosinteze. Plant Cell Environ. 27. 155-165 (2004).
Zhu, X.-G. Računarski Pristupi Guiding biotehnološkog poboljšanje usjeva fotosintetskom učinkovitost. Dr. teza, University of Illinois. Urbana, IL. (2004).
Zhu, X.-G., Govindjee Baker, NR, deSturler, E., Ort, DR, Long SP klorofila fluorescencije indukcijski kinetika u lišću predvidjeti model opisuje svaku diskretnu skup uzbude energije i prijenosa elektrona povezana s fotosustava II. Planta 223, 114-133 (2005).
Zhu, X.-G. de Sturler, E. Long SP Optimiziranje raspodjelu sredstava između enzima ugljičnog metabolizam može dramatično povećati stopu fotosinteze: numeričko simulacije pomoću evolucijski algoritam. Fiziologija bilja. 145, 513-526 (2007).
Zhu, X.-G., Duga, SP Ort, DR Koja je maksimalna učinkovitost s kojom fotosinteza može pretvoriti sunčevu energiju u biomasu?Trendovi u biotehnologiji 19, 153-159 (2008).
8/17/08
